by Eva Herbst, Structure & Motion Lab, The Royal Veterinary College, UK. If you would like to contribute a guest post, please get in touch, such as on Twitter or Facebook.

Fig. 1 Reconstruction of Crassigyrinus scoticus (Panchen & Smithson 1990)
My name is Eva Herbst and I started my PhD with John Hutchinson and co-supervisor Chris Richards this October 2016. I am researching limb and backbone anatomy in early tetrapods (four-limbed vertebrates), and their (lis)amphibian descendants, to find out more about the transition from water to land.
The first animal I am working with is Crassigyrinus scoticus, an aquatic tetrapod from the early Carboniferous (about 320 million years ago) of Scotland. Crassigyrinus has received the nickname “tadpole from hell” in our lab because it was up to two meters long and had huge eyes, tiny forelimbs, and large teeth (indicating predatory habits). In fact, the name Crassigyrinus comes from the Latin word ‘crassus’, meaning thick, and the Greek word ‘gyrinós’, meaning tadpole.
Crassigyrinus is remarkable not only for these features but also because it was clearly a completely aquatic animal; presumably more so than its older relatives Ichthyostega and Acanthostega. So this is an early example of a tetrapod that (probably) went entirely back to the water, with no abilities at all to walk, lumber or maybe even slither on land– but I’ll find out more about that!
Its skeleton also shows some other remarkable features. An interesting discovery made while investigating the Crassigyrinus fossil in the 1980s is three ribs that broke and then healed (Panchen 1985). How and why did these injuries occur? We don’t know yet.
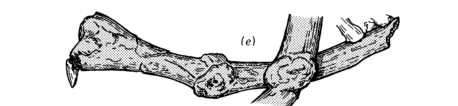
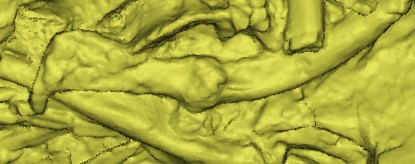
Fig. 2 Diagram of two healed ribs (Panchen 1985) above and 3D model of the bottom rib below.
The unusual round parts in the middle of the ribs are evidence of this repair process. Bone is able to remodel during an animal’s lifetime, even after the animal has stopped growing. New bone material can be deposited to repair a break in the bone, such as in these ribs.
The most striking feature of Crassigyrinus is its extremely small forelimb. The humerus (upper arm) is only about half as long as the eye socket is wide! The ulna (one of the two bones in your forearm) is also tiny: here it is in my hand so that you can get an idea of its size. Remember that this animal was up to 2 meters long, and I’m much shorter than that!

Fig. 3 Left ulna from R10000 (Eva Herbst)
To find out how such an enigmatic animal moved through the water, we are analysing its postcranial skeleton (everything behind the skull). Others (Dr. Laura Porro and colleagues) are studying its head and feeding habits.
The first discovered fossil that includes postcranial material is specimen R10000 from the Natural History Museum (London, UK). This skeleton is preserved in a slab of rock, with disconnected bones partially hidden by rock, scales, and other bones. We scanned the fossils with a microCT scanner to digitally separate individual bones without damaging the physical fossil.
Let’s take centrum 13 (a piece of the spine; backbone) as an example.

Fig. 4 The R10000 specimen, showing the position of centrum 13 (Panchen 1985)
Centrum 13 is partially submerged in the rock, so we can’t see all sides of it. It is also covered by two scales.

Fig. 5 Photograph of Centrum 13 of R10000 (Eva Herbst)
By scanning this area of the slab, I can virtually remove the bone from the surrounding rock via segmentation (see below, centrum 13 in turquoise).

Fig. 6 The modelling process: R10000 is shown at left (Panchen 1985), the slab section of interest in the center (Eva Herbst), and the CT scan model at right, with centrum 13 in turquoise
Segmentation is the process of making a 3D model from a stack of two-dimensional x-ray images. From these models, I can determine the structure of each bone. The shape and size of the contact surfaces between bones determines the range of motion of a joint. For example, contact surfaces between vertebrae determine the flexibility of the spine, and contact surfaces between limb bones determine the range of motion of the legs.
My goal is to create a complete model of the animal that can then be used to investigate how Crassigyrinus might have moved, and how well we can estimate that. Did it move by undulating its spine and tail (like an eel), or did it primarily use its hind limbs to paddle? How did these traits compare with those of other early or later tetrapods? I will investigate these questions by looking at the range of motion of the hind limb first, and later other parts of the body. Here is a similar study of the limbs of the early tetrapod Ichthyostega (Pierce et al. 2012).
I thank the Natural History Museum of London (Sandra Chapman) for access to the NHMUK specimen, and Professor Jenny Clack and Dr. Timothy Smithson at Cambridge for their discussions.
Further Resources:
More information on digital reconstruction methods:
Lautenschlager, S., 2016. Reconstructing the past: methods and techniques for the digital restoration of fossils. Royal Society Open Science, 3(160342).
Early tetrapod evolution:
Clack, J.A., 2012. Gaining ground: the origin and evolution of tetrapods. Indiana University Press.
References:
Panchen, A.L., 1985. On the amphibian Crassigyrinus scoticus watson from the Carboniferous of Scotland.
Panchen, A.L. & Smithson, T.R., 1990. The pelvic girdle and hind limb of Crassigyrinus scoticus (Lydekker) from the Scottish Carboniferous and the origin of the tetrapod pelvic skeleton. Earth and Environmental Science Transactions of the Royal Society of Edinburgh, 81(1), pp.31–44. Available at: http://dx.doi.org/10.1017/S0263593300005113.
Pierce, S.E., Clack, J.A. & Hutchinson, J.R., 2012. Three-dimensional limb joint mobility in the early tetrapod Ichthyostega. Nature, 486(7404), pp.523–526. Available at: http://dx.doi.org/10.1038/nature11124.

2 Comments Add yours